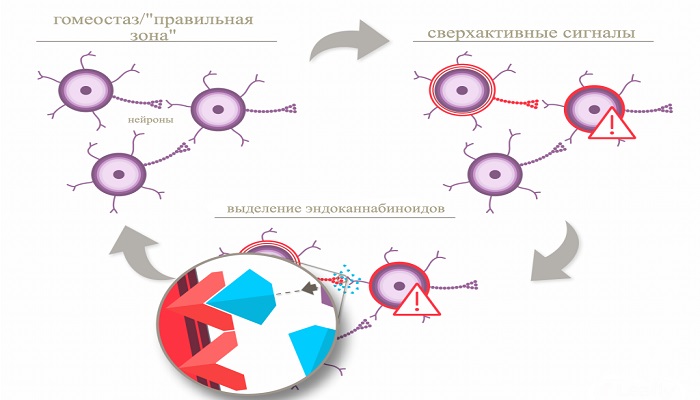
CB1R может обладать огромной сигнализационной сложностью. Самой простой функциональной единицей GPCR (в том числе CB1R), является гомодимер — это новобранцы гетеротримерного белка G. Тем не менее, CB1R физиологически «доминирует», поскольку он может расщепляться в Gi / O-белки даже в отсутствие агониста, тем самым истощая общий внутриклеточный Gi и ограничивая биологические сигналы других Gi / O-связанных рецепторов.
GPCRs также могут гетеродимеризоваться с другими рецепторами, что приводит к комбинаторному набору вторичных мессенджеров. Существуют доказательства сборки CB1R-μ-опиоидов, CB1-OX1 орексина, CB1-β2 адренергических, CB1R-D2 дофамина, CB1R-A2A аденозина и CB1R-тирозин-киназы B (TrkB) димеров рецепторов в частности.
С-конец CB1R содержит сайты фосфорилирования для GPCR киназы (GRKs), ведущие к β-аррестин-зависимым десенсибилизациям (АА 426/430), а также интернализации (AA460-473) домена. CB1R либо возвращаются в мембране либо помечаются GPCR связанными сортировками белка-1, что предназначено для протеасомной деградации.
AA465-473 оконечности CB1R могут связать адаптеры белков, таких как CB1R взаимодействующие белки 1а (CRIP1a) и 1b. Это последнее взаимодействие с CB1R может быть ограничено мозгом взрослого человека, потому что CRIP1a предназначен для соматодендритической области нейронов, и профиль экспрессии белка отличается от CB1R в мозге плода.
Сложность сигнализации на CB1R еще более усиливается эндогенными агонистами или антагонистами, синтез которых и деградация отличаются от эндоканнабиноидов.
Эти соединения могут модулировать наличие и сигнализацию компетенции CB1R. Гемопрессин является эндогенным нонапептидом, полученным из α-гемоглобина.
Хотя гемопрессин сам признается в качестве обратного агониста, продукты его усечения сообща функционируют как CB1R агонисты. Гемопрессин производно структурно и энергетически подходит лиганд-связывающему карману CB1R, влияет на CB1R и модулирует рост аксонов. Отсутствие гемопрессина вызывает эффекты в CB1R-/ — у мышей. Предположительно, гемопрессин может эффективно модулировать нейронные CB1Rs в естественных условиях.
Наконец, головным мозгом плода может быть воспринято быстро меняющаяся нейронная активность, как множество молекулярных переключателей управления в положении «включено» и «выключено» в точной последовательности. Материнское злоупотребление каннабисом может рассматриваться как патогенный раздражитель, провоцируя физиологический выход CB1R, тем самым перекрывая физиологический каскад сигналов эндоканнабисоленоидов.